Hynek's lab studies stem cells in hopes to better understand motor neuron development and diseases.
He began his talk introducing Hans Driesch's experiments with Sea Urchins in 1892. He, studying the embryo, separated the cells resulting after the first round of cell division. He imagined that the cells would then develop into the respective but separated portion of the sea urchin corresponding to the location where the cells were harvested. Instead each of the cells developed into fully functional complete sea urchins.
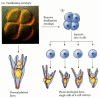
The initial cells Driesch harvested where embryonic stem cells, totipotent in nature, able to differentiate into any cell (not to be confused with: Pluripotent able to become cells that compose any of three germ layers, multipotent able to become many cells of a related family, Oligopotent differentiate into very few set of cells, and unipotent can only become themselves). So awesome, right? Let's just play with embryonic stem cells. But, really - let's!
In any developed organism the amount of cellular diversity, the systems of cross talk and proximity between these highly diversified cells makes studying specific cells very difficult. In vivo animal mutant models can be tricky and lead to embryonic lethality before gaining any new insight. Therefore, learning more about the genetic pathways and environments needed to induce stem cells to become specific types of cells can allow researchers to study those very cells more concretely. Wichterle aim's to do just that with motor neurons.
Hynek continues on to point out that due to the diligence of labs previous research (please visualize the head nod that ensued in the direction of his previous mentor, Tom Jessell) we know ways to distinguish motor neurons based on molecular identity and functionally. The signals required to develop spinal cord motor neurons are also being elucidated: starting from the rostral or cervical end of the spinal cord (the neck) retinoic acid (RA) signals at its strongest then lessens as a gradient down the spine through the brachial region towards the middle of the cord meeting the least of the increasing gradients of GDF, FGF and WNT (signaling morphogenic molecules).

Wnt, GDF, and FDF increase along the thoracic level till reaching the end of the lumbar or caudal spinal cord. Lastly, across the entire ventral portion of the spinal cord, SHH signals as a gradient strongest from ventral lessening upwards into the dorsal (top half) area of the cord.
Using these tools and signals, the Wichterle lab has been successful in inducing pluripotent stem cells in vitro to differentiate into cervical and brachial motor neurons. They express the appropriate markers, when injected back into a developing spinal cord they migrate to the correct distinct positions, and can then function perfectly as the in vivo developed cells within the tissue. Now with the model cells to work with the lab moves on to test the role of repressive chromatin in the pattern of developing motor neurons in the rostral to caudal axis, microRNA's in the dorsal-ventral axis, and modeling neuronal diseases like ALS.
Hox gene products, homeobox proteins, are an important family of transcription factors during development; they aid in the creation of different motor neuron classes across the Rosto-Cadual axis (length from the neck to tail). Different sets of Hox genes are required to be expressed in different anatomical locales along this axis. For example, starting Rostal, the cervical region requires Hoxc genes 4-5, Brachial requires Hoxc6-8, and (moving caudual) Thoracic Hoxc9 and Lumbar Hoxc10-13.

The hoxc genes are interestingly assembled in the genome on chromosome 12 in a similar fashion to expression; hoxc gene 4-5 are organized in the 3' end in chromatin (DNA) and increase in order of number till the 5' end. Therefore, it is thought there must be repression of specific hox expression in these distinct anatomical locations to allow for the differential gene expression (i.e. repression of hoxc4 & 5 in the lumbar section but none of hoxc10-13 permitting their expression). It was postulated through correlations that during development repression is lifted from the primary hoxc genes, priming the Chromatin structurally, allowing transcriptional molecular machinery access for the later relaxation (de-repression) of the numerically higher hoxc genes. Therefore, it was postulated that there is a progressive clearance of repressors on the chromatin starting with Hoxc4 and continuing to Hox13 during the due time of need for induction of those cells.
Wichterle and Dansen labs collaborated (NYU, NOTE: post-doc from Jessell lab as well) to confirm the postulated nature of the Hoxc chromatin. They performed chromatin immunoprecipitations on stem cells and found that all hox genes had a considerable amount of repressive markers across them. They looked at the different cell types in the cord and found in development cervical motor neurons lack of repressive markers. Puzzling, they decided to do Quantitative PCR to profile the hoxc gene expression during time points in the developmental cascade of inducing stem cells to motor neurons. They found that there was no progressive clearance of repression, the profile of hox gene repression did not correlate with expression. They decided to do more chromatin immunoprecipitations but looked for the density of retinoic acid receptors. They found a correlation between the presence of RA receptors located on the hoxc 4-5 genes and the loss of repressive markers. Wichterle states that somehow the recruitment of RA receptors initiates the clearance of repression. Cool, How?
Its pretty convenient that the two most well know players in the patterning of motor neurons along this axis show tight correlation between their activities. Can you further confirm RA receptors role by stunting it's gradient or prolonging its gradient along the axis, then look at methylation (this is the marker they used for repression, methyl groups added on chromatin usually mark a repressed gene)? That is, at a time when the changing RA gradient won't completely mess up the fate of the cells...?
What are the molecular complexes that are responsible for providing the methylation on the chromatin? What is the gene expression profile for these chromatin remodelers - Micrroarray and look for enrichment of certain known chromatin modifiers?
Wichterle goes on to question how the caudal neurons develop. It seems throughout the development of the cells there is no lack of repressive markers. They looked at a microarray to see what genes were enriched in the lumbar cells versus the cervical. They found one interesting gene, Cdx2. Inducing stem cells with cdx2 seemed to have no role, but they found in cooperation with FGF they could induce expression of hoxc8 motor neurons. Through further experimenting they found they could also induce hoxc6-9 genes but repress hoxc1-4 genes. He concludes showing that hoxc9 likely cooperates in the repression of hoxc1-4. (For more information see paper)
The orchestra is getting meatier regarding the instruments to play out development of even one cell type: Signals, transcription factors and chromatin modifiers...
Sorry for the length of this one...
Hynek continues with part two, what I found to be the most interesting data, about the dorso-ventral patterning of the spinal cord. SHH signals as a gradient from the notocord and floorplate (Ventral/bottom of Spinal cord aka the side facing the front of your body) which is responsible for inducing different varieties of Interneurons and Motor neurons based on the receieved amount of SHH. In the ventral portion, closest to the floor plate, cells are induced to express NKx2.2 as the gradient lessens dorsally the next class of express Olig2 (Motor neurons) and then further lessening of the gradient the next class express Irx3 (picture: http://en.wikipedia.org/wiki/File:Sonichedgehog.jpg). These genes are markers of progenitor cells, the cells expressing them have matured from stem cells and based on gene expression decided to be a specific type of cell. (ex. nkx2.2-motor neurons, olig2-oligodendrocytes).
Hynek's lab investigated the role of microRNAs in cells that border an area with overlapping gradient signals.
Presumably these two gradients are providing the cells with signals that instruct competing progenitor gene expression to either become one type of cell or another. Somehow cells naturally overcome this confusion and the spinal cord results in a clear pattern of segregated neuron types.
The lab noticed previous research done that identified a gene that when knocked out would eliminate all microRNAs, Dicer. Dicer mutants do not live past gastrulation, therefore studying it's affects have been proven difficult. Wichterle's lab formulated a conditional tamoxifen-inducible line for dicer. Allowing them to knockout dicer's expression during specific time frames in development. They still had issues with survival of the animals but had enough success to illustrate an expansion of motor neurons and losses of Irx3 cells. Seemingly, in the natural background microRNA represses the expression of Olig2 allowing only a discrete segment of the spinal cord to develop into motor neurons. This indicates that the p2 set of cells (Irx3 expressing) must transiently express olig2 at some point. The further investigated the situation by looking at the microRNA profile of cells exposed to high vs. low levels of SHH.

To Be Continued...
This is very long. I do apologize. I think I just got really excited about the research...my head is exploding with information.
No comments:
Post a Comment