Expression of major guidance receptors is differentially regulated in spinal commissural neurons transfated by mammalian Barh genes.
Developmental Biology
Kawauchi, Muroyama, Sato, and Saito
The authors claim that the competence of spinal cord neurons to send axons ventrally across the midline to become commissural neurons is temporally dependent on embryonic stage. They focused their studies on the known guidance receptors and transcription molecules that mark and aid Math-1 derived D1 interneurons in their characteristic maturation. During said embryonic stage misexpression of mbh genes activates guidance receptor genes, Rig1 and Np2. Lhx2 has been shown in our lab to have a strong potential genetic relationship with Rig-1, furthermore, Lhx2 mutants do not extend their axons through the midline, instead their trajectory stops adjacent to the floor plate in the ventral spinal cord. Kawauchi claims to have revealed that mbh genes (mbh1 and mbh2) control pathways either lhx2-dependent or independent of guidance receptor expression.
I was under the impression this information was already known…
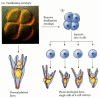
Kawauchi introduces the basics of commissural neuron phenotypes in the developing spinal cord:
The transient and simultaneous expression of chemoattractants and receptors, as well as, chemorepellents and their receptors is a very intriguing display of the amazing molecular circus. Expression and function of these groups must be temporally and distinctively expressed locations allows cells types to obtain their distinguishing properties for a functioning growing adult.
Ignoring the Roof plate for the moment, we know that attractants, like netrins and sonic hedgehog, and repellents, like slits and semaphorins, are produced from the floor plate acting upon commissural neurons aiding in their trajectories. Some of the key chemo-guidance molecule/receptor pairings attracting to and across the floor plate are: Netrin-Dcc, Sonic hedgehog-Boc, Slit–Robo1, and Semaphorin-Npr2.
So please imagine this for a moment: tons of neuronal cells are being born and oozing out of the ventricular zone in the spinal cord. They have been seemingly fated by their progenitor gene expression to begin a cascade of further gene expression requiring them pretty much instantly to migrate their cell bodies to a particular position and simultaneous send their axons to specific locals. Numerous cells are engaged in this activity at the same time but we end up with very diverse groups owing to evolutionarily distinct fully functioning organisms. While in this discussion we are concerning ourselves with the understanding of a specific cell type and genetic pathway, if you think of multiplying these ideas by the millions, the image of orchestration of development becomes a mystifying tale. (A tale that always leaves me hungry and smiling).
Focusing on Rig-1 (Robo3), it functions to repress the role of the Robo1 receptor allowing cells expressing Rig-1 to ignore Slit repulsion until after crossing the floor plate. Robo1 is then used to suppress Dcc to prevent re-crossing of the axon through the floor plate (Plus, it has also been shown with growth cone studies that Slit’s repulsion dominates Netrin’s attraction). Kawauchi et al, go on to investigate the transcriptional relationship between Lhx2, Rig-1, and the bar-class homeobox genes mbh1 (barhl2) and mbh2 in fating the commissural Math1derived dorsal spinal neurons.
After in vivo electroporation of mouse embryos at E11.5 & E12.5 with Mbh1, Mbh2, Lhx2, Lhx9, and Math1 they provided some evidence supporting the bar-class genes playing an important role between Math1 expression and the Lim HD transcription factors.
One first notices d1 neurons by the expression of Math1. Math1 is a neuronal progenitor solely responsible for birthing all commissural projecting Lhx2+/Lhx9+ cells in the dorsal most area of the spinal cord beginning at E9.5. Once cells have turned off Math1 they, based on embryonic day, express Lhx2 and Lhx9 (more specifically, in my studies they express them temporally not only based on their dorsal-ventral migration location but also based on their RostroCaudal location). But the genetic-molecular control of this transcriptional control is not well understood.
They found that ‘misexpression’ of mbh1 and mbh2, but not Lhx2, by transfection into right side of the spinal cord at E11.5 (then allowed expression for 2 days till analysis at E13.5) have almost redundant roles in providing an >10% increase in commissural projecting axons. (*Of note to me, there are seemingly some ipsilateral projecting cells labeled on the medial lateral edges). Transfection at E12.5 analysis at E14.5 does not provide the same effect and cells obtain a phenotype copying transfection of Fluorescent reporter DNA only.
(*I would like to note that at this point in the paper there is a shift of focus. In my opinion this shift is modestly expressed. If I were writing the paper I would have tried to highlight the importance of the information/analytical ideas expressed. I understand that the shift is based on theoretical ideas - that they only support these ideas they do not concretely provide evidence. But it lays down, to me, a good path for investigation).
This is exciting -maybe. As in, the cells are not as malleable to gene expression instructions that they may have been a day or few earlier. Have they past some threshold in development where new waves of genes have been expressed to allow a suppression of the old ones (similar to the Rig1/Robo1/Dcc story)?? The authors say the cells transfected at E12.5 are not ‘competent’ to become commissural neurons.
ORRRR (bear with me) have a certain amount and type of genes relating to specific functions of that neuronal cell already been expressed and sequestered to specific areas in the cell, so, when they are needed (as RNAs with the machinery available to express them as functioning proteins when signaled to be needed) to respond to the transient distinct spatial signals from the environment they have a stock to do so efficiently.??
Kawauchi again transfects cells at E11.5 with mbh1 and mbh2 then examined changes in gene expression for most of the known guidance receptors. Robo1, Robo2, Kit, Boc, and Dscam were not activated by misexpression of either Bar-class genes. However, Rig-1 and Nrp-2 were both elevated. Nrp-2 elevated expression does not change 2 days after transfection when the cells have reached the deep dorsal horn settling position (mirroring the endogenous expression). However, Rig-1 expression remarkably disappears by the time the cells reach the deep dorsal horn 2 days after transfection. The authors imply that this indicates that Rig-1’s activated misexpression by mbh is able to be saved/regulated by innate pathways naturally downregulating Rig-1 at the appropriate time/settling position. Mbh1 did not activate Mbh2 and mbh2 did not activate mbh1. Transfection with Lhx2 activated Rig1 but Lhx9 transfection did not (both Lhx gene tranfsections had no activating affects on Mbh1 or 2).
Interestingly, Mbh1 and 2 transfection has been shown to increase Dcc protein levels. However the misexpression does not result in an increase/activation of Dcc gene transcripts, suggesting an increase in Dcc protein is based on activating post-transcriptional expression as the authors state. AWESOME.
Math1 transfection into the spinal cord activated Mbh1, Mbh 2 , Lhx2, and Lhx9 gene expression. (Note: Lhx9 activation was weaker by Mbh genes than my Math1 suggesting there are more Mbh-independent factors involved or a timing issue). To decipher the order of gene expression, since Mbh1 and Mbh2 also activate Lhx2/9 genes, Kawauchi transfected Math1 and a construct that expresses dominant negative form of either Mbh1 or Mbh2. They found the activation effect on Lhx2 and Lhx9 genes by Math1 was repressed in the presence of the inactive form of Mbh1. But Mbh2 activation by Math1 was not repressed in the presence of the dominant negative Mbh1 proteins. (Note: repression of Lhx9 was not complete therefore more of an indication that there are mbh-independent pathways leading to lhx9 expression).
They found that it is the repressive abilities of Mbh1 and Mbh2 (through expression of chimeric Mbh proteins consisting only of the homeodomain and transcriptional repressor domain) that allow for activation of Lhx2 and Lhx9, therefore, most likely activation is an indirect effect.
Further strongly supporting the idea that there is another in-between factor from Mbh’s to Lhx2/9.
Well I guess we can somewhat confidently say that between Math1 and Lhx2/9 Mbh1 and Mbh2 repress expression/activity of another factor that represses Lhx2/9 therefore aiding in their expression.
I think animal models with single or combinational mutant knockouts might provide more evidence.
I wonder if there have been studies involving axon severing then analyzing the ability of the growth cones to express/perform different proteins/functions in response to different signals (or move…probably would definitely not be able to move…but maybe reorient)...
I'm thinking that if mRNA transcripts are produced and localized to specific areas in the axon or growth cone AND if the cell has produced transcripts but not the protein - The cell may be waiting for further signal to express the protein, therefore, if they are pre-produced and localized based on the cell type (mutant background, etc.) we can make some comments on how the severed growth cone acts in response to extracellular signals.
Just brainstorming...

 (Or really detect anything at all - the literature and in situ's done suggest that Math1 expression in the spinal cord begins at E9.5 and continues till E13ish. Therefore, my tamoxifen treatment may not have induced any GFP expression OR dependent on how long the tamoxifen stays active in the system it may induce and label the first round of cells to differentiate into the D1 class.)
(Or really detect anything at all - the literature and in situ's done suggest that Math1 expression in the spinal cord begins at E9.5 and continues till E13ish. Therefore, my tamoxifen treatment may not have induced any GFP expression OR dependent on how long the tamoxifen stays active in the system it may induce and label the first round of cells to differentiate into the D1 class.)