As a young scientist, especially at Columbia, I am taught (and heard the complaints from Grad students) that one should be going to every seminar, doing back-to-back experiments, refreshing pubmed search every hour, and reading papers in every spare minute. Those who succeed are consumed with this process and sometimes smell bad.

While I am consumed by this lifestyle and am ready to throw myself completely into a project - I am scared. If I throw myself into my work completely I'm afraid I'll lose everything else about myself. I want it all. I don't want to be like everyone else and I don't think I have to be. I just wish people didn't judge you for not filling and/or following the footprints perfectly.
Science: I've just finished reading Motor and Dorsal Root Ganglion Axons Serve as Choice Points for the Ipsilateral Turning of dI3 Axons by Oshri Avraham,1 Yoav Hadas,1 Lilach Vald,1 Seulgi Hong,2 Mi-Ryoung Song,2 and Avihu Klar1 published in the Journal of Neuroscience. This entry will discuss the study.

This image depicts the six classes of dorsal interneurons to arise during development in the spinal cord (Embryonic days E9.5 to E13.s in mice) adjacent to the roof plate. From the roof plate a gradient mainly of BMPs and Wnts influence newly born cells based on time of birth and position to express a specific neural progenitor gene. Each of the six classes are produced by a unique neural progenitor and can be identified by a unique combination of known transcription factors, their settling pattern and axon trajectory. In my lab we mainly focus on the D1 class of interneurons (and I primarily am focused on examining further genes/gene expression patterns that which instruct these cells to mature into their less characterized sub populations and eventually use our mouse model to uncover these cells contributions to the adult somatosensory networks).
In this paper, the authors examine the axon trajectory of the D3 interneurons.

Interneuron development/patterning in the spinal cord begins around E9.0 by various gradient produced cocktails of morphogens inducing expression of neuronal progenitors. The progenitors in turn promote the expression of various BarhL and LimHD proteins that not only help identify specific neuronal types but also seem to direct axonal trajectory based on their levels of expression in the cell (Wilson et al. 2008)(However, little is known regarding the targets and/or the mechanisms by which these proteins function).
The path of axon projections is said to be governed by the growth cone interactions with various immediate cues (guiding them to turn their terminals in different directions) and long range cues (repulsion from the roof plate/attraction by the floor plate/later repulsion by the floor plate after crossing). This paper proposes that at least one group, d3, uses architecture and possibly, Isl1 (cell-cell interaction), to guide them in their turning decisions when projecting laterally out from the spinal cord to their final targets.


First the authors sought to characterize the population of cells in more detail. We know that these cells express Tlx3,Prrxl1, Brn3a, Olig3, Asc1 and they are the only interneurons to express Isl1 (which is also expressed in Motor and Dorsal Root Neurons). They searched for evolutionary conserved non-coding enhancer elements around the Isl1 gene via Enhancer Browser Project. They cloned three different regions fused CRE recombinase downstream of the enhancer and screened for labeling by electroplating Chick neural tubes along with a conditional nGFP reporter vector. They found a combination of using two elements specially labeled the dI3 cells (referred to as: EdI3). They subsequently used this system to label and observe the intrinsic nature of the cells.
{kind=link}
Based on axon projection and turning point for lateral growth they defined four distinct populations of DI3 neurons: Dorsal projecting turning caudal or rostral at dorsal funiculus, Ventral projecting turning caudal or rostral at ventral lateral funiculus. The ventral projecting axons tend to turn/follow the motor neuron axons out of the ventral spinal cord and the dorsal projecting neurons follow the DRG neurons out of the dorsal cord to then turn laterally towards final targets.
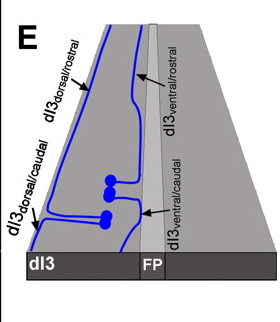

The top figure above illustrates the four types of Di3 neurons identified as observed in 'open book presentation'. Basically they took the roundish spinal cord and cut down the midline from roof plate to the floor plate and opened the cord like a book. See illustrated example below from Dickson BJ, Gilestro GF (2006) Regulation of commissural axon pathfinding by slit and its robo receptors. Annu Rev Cell Dev Biol 22: 651–675.

The authors further state that the axons of the dorsal interneuron classes do not intermingle, their cell soma positions get more mixed between each class as they migrate ventral or dorsal but their longitudinally projecting axons bundle together specific to their class.
They go on to investigate the molecular actions of Isl1. The LimHD genes have previously been shown to play no role in cell fate (Avraham et al., 2010; Kania and Jessell, 2003; Pillai et al., 2007; Luria et al., 2008; Wilson et al., 2008) but they are emerging to have roles in maintaining a cells fate and their axon patterns. Avraham electroporated Chick neural tubes at HH14 (premitotic) and HH19 (postmitotic) with nGFP and Lhx9 or Isl1 then quantified the protein expression of the limHD proteins compared to the control (un-electroporated side).
 They found that there is a cross-repressive relationship between Lhx9 and Isl1 in Pre- and Postmitotic cells. Aberrant expression of Lhx9 imparts a reduction in Isl1 protein as compared to the controls. A similar decrease in Lhx9 protein is observed when electroporated with Isl1. They also tested and found cross-repression between Lhx1 and Isl1.
They found that there is a cross-repressive relationship between Lhx9 and Isl1 in Pre- and Postmitotic cells. Aberrant expression of Lhx9 imparts a reduction in Isl1 protein as compared to the controls. A similar decrease in Lhx9 protein is observed when electroporated with Isl1. They also tested and found cross-repression between Lhx1 and Isl1.The expression patterns of other transcription factors known to mark these cells are not changed (Brn3a, Pax2, Tlx3). Avraham concludes that ectopic expression of Isl1 does not affect the fate of other cells and does not induce a population increase of dI3 neurons. He supports his conclusion by taking a look Isl1hypo- mutant mouse. In this mouse line the motor neurons express some Isl1 but the dI3 neurons are lacking Isl1 protein completely. So the dI3 cells are generated even in the absence of Isl1 - they just must not project correctly.
Next they directed ectopic expression of Isl1 in dI1 class neurons in the Chick cord (they used the specific enhancer element expressed in dI1 neurons with CRE fused downstream and electroporated the construct with a Lox'd Isl1 taumyc tagged expression construct) to examine if expression of the Isl1 gene would confer a dI3-like axonal projection pattern on the dI1 neurons.
The dI1 neurons send axons ventrally towards the floor plate and cross the midline turning in the ventral lateral funiculus (contralateral/commissural) but they also send axons ipsilaterally (same side) in the medial towards ventral lateral funiculus. While some dI3 neurons project dorsally none of the dI1 neurons send their axons in the dorsal direction. The authors state about 20% of dI1 neurons project to the contralateral side but in the dI1-Isl1 expressing neurons only 10% cross the midline. They observed these neurons to turn laterally upon reaching the motor neuron zone, follow their tracks (like dI3-ventral neurons) and turn laterally. They found no change in the dI1-ipsilateral-Isl1 expressing neurons.

Is this due to timing of expression, Higher Lhx9 expression in the ipsilateral population (Wilson 2008) allowing for functional repression of Isl1, or (as the Authors discuss) the axons of the dI1-ipsi never get close enough to the motor neurons?
They discuss that Isl1 may infer a short range adhesive interaction between the motor neurons and dI3 neurons that is required for their turning lateral. They do state that this can be challenged by manipulating the motor neurons/DRG neurons.
I think they should have also targeted Isl1 reduction in motor neurons. If you knock down Isl1 in the motor neurons - do the ventral projecting dI3 neurons or dI1-contra-isl1 expressing neurons still turn laterally upon reaching them?
Although LimHD protein repression was illustrated as a molecular consequence of Isl1 expression, I wonder what the other targets of Isl1 are (and the other LimHD proteins specific to the other dorsal classes). There must be other players in the scheme that are repressed or promoted....
No comments:
Post a Comment